A nitrogén-, a foszfor- és a káliumtartalmú vegyületek, legtöbbször vízben nem oldható növényi részekben, a sejtfalban találhatók nagyobb mennyiségben. A mikrobák a növényi anyagok bontása során adszorbeálják és abszorbeálják a felszabadult kationokat és anionokat, azokat a talajba juttatják, ahonnan a növények számára újra felhasználhatóvá válnak.
![]()
A maradványokat bontó mikroorganizmusok anyagcsere-intenzitása mindig az adott ökológiai feltételekhez (hőmérséklet, nedvesség, kémhatás, egyéb anyagok jelenléte) alkalmazkodik, és attól függően mindenkor maximális sebességű. A baktériumok és a gombák hatékonyan ásványosítják a nem vízoldható növényi maradványokat a morfológiai tulajdonságaik, a növekedési és a szaporodási tulajdonságaikból is adódó enzimatikus képességeik miatt. A maradványokon megtapadó mikrobák közvetlen környezetükbe választják ki a termelt enzimeiket, ezért hatásuk pontszerű.
A szerves anyag apró darabokra töredezik és a cellulózbontó mikroorganizmusok együttműködve az ott jelenlévő egyéb nem cellulózbontó mikroorganizmusokkal, sokkal intenzívebb cellulózbontásra képesek, mint önmagukban. A búzaszalma összetétele az említett elemeken felül 34-40 % cellulóz, 30-35 % hemicellulóz és 14-15 % lignin, amelyek a sejtfalban bonyolult szerkezetet alkotnak.

A mikrobák a növényi maradványok bontása során a növények számára újra felhasználható tápanyagokat szabadítanak fel
A Cellulóz
A növényi biomassza legnagyobb hányadát, mintegy 40%-át a cellulóz polimerek alkotják. A cellulóz homopoliszaharid, és a legkisebb ismétlődő egysége a cellobióz, mely két β-1,4-kötéssel kapcsolódó glükóz molekulából tevődik össze, és benne a glükóz monomerek egymáshoz képest 180°-kal elforgatva helyezkednek el. A cellulóz hosszú (10 000-15 000 glükóz egységekből áll) lineáris polimer, mely inter- és intramolekuláris hidrogén kötések kialakítására nagymértékben hajlamos, az így stabilizált makromolekula nagyfokú rendezettséggel jellemezhető. Vízben oldhatatlan, kémiailag stabil és enzimeknek is viszonylag ellenálló. Az elsődleges sejtfalban a cellulóz rostok átmérője jellemzően 3 nm. A cellulózt a roston belül kristályos és amorf (parakristályos) részek alkotják.
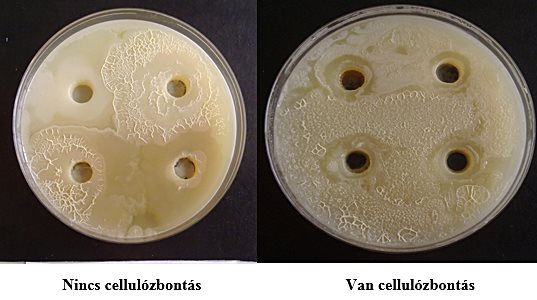
A cellulózbontó törzsek vizsgálatára a Fuller-Normann-féle cellulóz-dextrin és karboximetil-cellulóz tartalmú agarlemezek alkalmasak. A cellulózbontás mértékét a lyukak körül kialakult degradálódott gyűrű mérete és erélye mutatja.
A hemicellulózok
Az évelő és az egynyári növényekben a hemicellulózok alkotják a biomassza 25-35%-át. A hemicellulózok mindazon nem keményítő jellegű poliszaharidok, amelyek a magasabb rendű növények elsődleges és másodlagos sejtfalában a cellulózzal asszociálva fordulnak elő és lúggal kivonhatók, feltárhatók. Polimerizációs fok és összetétel tekintetében nagy változatosságot mutatnak. A lánchosszúságuk általában kisebb, mint cellulóz esetében, ugyanakkor nagyszámú oldallánc helyezkedik el a fő vázon, így a hemicellulózok a cellulózhoz viszonyítva kevésbé kristályos, reakcióképesebb molekulák. Leggyakoribb vegyülete a xilán. Legfőbb alkotóelemeik az L-arabinóz, a D-galaktóz, a D-glükóz, a D-glükuronsav, a D-mannóz és a D-xilóz, de ezen kívül L-frukóz, L-ramnóz és L-galaktóz is megtalálható oldalláncaikban. A hemicellulózok a cellulóz láncokat beburkolva védik, illetve összeköttetést biztosítanak a cellulóz láncok és a lignin régiói között.

A növényi maradványok mikrobiológiai lebontása a humuszképződés és a kórokozó gombák visszaszorítása miatt is fontos
A baktériumok növényi maradványokat bontó enzimei
Az oldhatatlan cellulóz hidrolízisének, és hasznosításának érdekében a cellulóz bontására képes mikroorganizmusok szabad, vagy sejthez kötött extracelluláris celluláz enzimeket termelnek. Az enzimaktivitások három fő típusa található meg a teljes celluláz enzimrendszerben:
- (a) az endoglukanázok,
- (b) az exoglukanázok,
- (c) a β-glükozidázok.
Az endoglükanázok a cellulózláncból hasítanak változó hosszúságú oligoszacharidokat, és ezzel együtt új láncvégeket hoznak létre. Az exoglükanázok a cellulóz redukáló és nem redukáló végeinek hasításával glükózt, vagy cellobiózt szabadítanak fel. Az amorf cellulóz részen támadni képes endo (belső)-1,4-β-D-glükanázok a cellulózt véletlenszerűen hasítják, hosszabb-rövidebb kisebb, de mindenképpen rövidebb hosszúságú láncokra. Ezen cellulóz láncok végéről a kristályos cellulóz hasítására is képes cellobio-hidrolázok cellobiózt hasítanak le, melynek további hidrolízisét az 1,4-β-D-glükozidázok, más néven cellobiázok katalizálják.

A búzaszalma degradálódása 2 hónap (fenti két kép) és 4 hónap (alsó két kép) alatt cellulóz bontására képes Bacillus sp. és Pseudomonas sp. törzskeverékek felhasználásakor szabadföldi vizsgálatokban.
Az endoglükanázok a cellulóz poliszacharid lánc amorf régiójában hasítanak, mellyel változó hosszúságú oligoszacharidokat, és ezzel együtt új láncvégeket hoznak létre. Az exoglükanázok a cellulóz redukáló és nem redukáló végeinek hasításával glükózt vagy cellobiózt szabadítanak fel. A β-glükozidázok a cellobióz glükózzá történő hasítását végzik. A β-glükozidázok baktériumokban és gombákban a celluláz enzimrendszer fontos részét képezik, ahol a rövidebb szálú oligoszacharidok és a cellobióz glükózzá történő hidrolíziséért felelősek.

A gabona- és repcetarló szakszerű kezeléséhez hozzátartozik a növényi maradványok lebontását segítő mikrobiológiai készítmények használata
A cellulóz a Föld legelterjedtebb makromolekulája, mely a növényi sejtfal alkotóelemeként a leggyakoribb összetevője a növényi biomasszának. A cellulóz a növényi szárazanyag tömegben kb. 35 - 50% koncentrációban található meg. A természetben a cellulóz csak ritkán fordul elő tiszta állapotban, a legtöbb esetben egyéb vegyületek pl. hemicellulóz és lignin közé ágyazva található. A lineáris cellulózban a glükóz molekulák β-(1→4) glikozidos kötéssel kapcsolódnak egymáshoz.
A cellulóz rostok hemicellulózokkal borítottak, melyekhez lignin régiók kapcsolódnak, emiatt korlátozott a cellulázok hozzáférése a cellulóz szálakhoz. Amennyiben a rendszerben (talajban) un. segítő enzimek is képződnek, a növényi maradványok bontása kedvezőbb lehet és rövidebb lánchosszúságú molekulák keletkeznek. Az ekkor képződő enzimek lehetnek a hemicellulázok, a lignint bontó mangán-peroxidázok, lakkázok (rezet tartalmazó oxidáz enzimek) és lignin-peroxidázok, valamint a pektinbontó enzimek.
A másodlagos sejtfal fő hemicellulózának, a hidrolízisében endo- és exoenzimek együttesen vesznek részt. Az endo-1,4-β-xilanázok az oldhatatlan xilán láncot rövidebb, oldható láncokká és xilobiózzá (két egységből álló molekula) hidrolizálják, amit az 1,4-β−D-xilozidázok xilózzá alakítanak. A xilán láncok gyakran tartalmaznak a láncon egyéb szénhidrátokat, amelyeket újabb enzimek hasítanak le. További savcsoportok is lehetnek a hemicellulóz láncon (pl. ecetsav), ez a növény fajtájától függő sajátosság.
Összefoglalóan megállapíthatjuk, hogy a holt szerves anyag bontása bonyolult mikrobiológiai, biokémiai és mikrobiális ökológiai folyamatok eredményeként jellemezhető. A hatékony szárbontás érdekében a gabona- és repcetarló szakszerű kezeléséhez hozzátartozik a növényi maradványok lebontását segítő mikrobiológiai készítmények hatásainak ismerete és használata.
Forrás: Balázsy Sándor, Mikroorganizmusok a növények szolgálatában. 2015, pp. 156, Kézirat






_fill_100x75_0.jpg)








_fill_99x55_0.jpg)



